Отдел Хвощевидные. Общая биологическая характеристика, классификация, значение
Растения, входящие в отдел хвощевидных (Equisetophyta ), отличаются от других споровых побегами, составленными отдельными члениками (междоузлиями) и узлами с мутовчато расположенными листьями. Характерной чертой хвощевидных является также наличие у них спорангиофоров – особых структур, несущих спорангии. Мутовки спорангиофоров образуют на стебле спороносные зоны, чередующиеся с обычными вегетативными листьями, или сидят на концах осей, образуя стробилы.
Ископаемые остатки хвощевидных известны из девонских отложений. К концу этого периода хвощевидные уже прошли длительный путь эволюции, а расцвет этой группы растений пришелся на каменноугольное время. В палеозое хвощевидные отличались исключительным разнообразием: среди них встречались травы, кустарники, лианы и деревья, достигавшие 20 м высоты. Вместе с гигантскими плауновидными эти формы образовывали каменноугольные леса.
В то время, помимо представителей класса хвощевых (Equisetopsida ), на Земле произрастали и хвощевидные растения другого класса – клинолистовых (Sphenophyllopsida )1 . Клинолисты (род Sphenophyllum ) образовывали густые заросли в лесах каменноугольного периода (рис. 1). Побеги некоторых растений этой группы достигали в длину нескольких метров при очень небольшом диаметре (0,1–0,5 см) и были либо полегающими, либо лазающими, о чем свидетельствуют прицепки, обнаруженные на стеблях одного из видов. К узлам молодых участков стебля прикреплялись мутовки из 6–9 клиновидных листьев.

1,2 – клинолист клинолистный (Sphenophyllum cuneifolium) (1 – общий вид части растения, 2 – листья разных узлов стебля); 3–5 – стробилы представителей порядка клинолистовых
(3 – сфенофиллостахис водяной (Sphenophyllostachys aquensis ); 4–5 – сфенофиллостахис Досона (S. dawsonii ), (4 – продольный срез части стробила, 5 – поперечный срез стробила в области междоузлия; сп – спорангий, сс – стерильный сегмент спорангиофора, вс – веточка спороносного сегмента спорангиофора)
Древние представители класса хвощевых – каламиты (Calamites ) – в каменноугольном периоде в значительной мере составляли древесный ярус заболоченных тропических лесов. По внешнему виду каламиты напоминали современные хвощи, только гигантских размеров (рис. 2). Стволы каламитов достигали 10–20 м высоты и 0,5 м в диаметре за счет способности к вторичному утолщению (слой вторичной древесины в их стеблях достигал 12 см).
1–3 – каламиты (Calamites) с разными типами ветвления; 4 – листья каламитов; 5 – поперечный срез стробила каламита; 6 – часть стробила, вид сбоку: слева – с поверхности, справа – в продольном разрезе (б – брактея, пп – проводящий пучок, сф – спорангиофор, сп – спорангий)
Листья, основные фотосинтезирующие органы каламитов, длиной от 2 до 7 см, были расположены в узлах стебля мутовками по 3–70 штук. Стробилы, длиной около 12 и шириной около 4 см, состояли из чередующихся мутовок стерильных листьев и спорангиофоров. Возможно, все каламиты были разноспоровыми растениями, т.е. формировали мужские и женские споры, а на образовывавшихся из них гаметофитах (заростках), соответственно, развивались либо только мужские, либо только женские половые клетки. У одного из каламитов (каламокарпон замечательный – Calamocarpon insignis ) найден мегастробил, в спорангии которого развивалась всего одна мегаспора. Она прорастала прямо внутри спорангия, образуя женский гаметофит. Такая крайняя степень редукции числа мегаспор в спорангии отмечена и у древовидных плауновидных палеозоя 2 .
В отличие от большинства древовидных плауновидных и класса клиноситовых каламиты пережили каменноугольный период и были обильно представлены и в пермское время. Но к началу мезозойской эры каламитовые, достигшие высокой степени специализации, вымерли под воздействием резкого изменения климата. Их менее специализированные сородичи из класса хвощовых на некоторое время пережили климатические перемены и в первой половине мезозоя в некоторых областях Земли даже образовали почти чистые сообщества по берегам водоемов. Однако и они постепенно почти все вымерли, так что в наше время весь отдел хвощевидных представлен одним родом хвощ (Equisetum ), относящимся к классу хвощовых и включающим около 20 видов.
Латинское название Equisetum впервые употребил Плиний Старший, имея в виду, по-видимому, сходство ветвистых побегов хвоща с хвостом лошади (от лат. equius – лошадь и saeta, seta – щетина, жесткие волосы). В русском названии «хвощ» отражено сходство взрослого растения с хвостом или с пучком волос. В некоторых местностях России хвощи с мутовчато расположенными побегами называют «сосенка», «елка», а по наличию у некоторых видов бурых спороносных побегов, похожих на пест, – «пестиками» или «толкачиками».

1 – участки горизонтального и вертикального корневища с клубнями у хвоща полевого; 2 – то же у хвоща болотного; 3 – схема поперечного среза клубня хвоща полевого; 4 – схема поперечного среза корня хвоща полевого (вп – воздушная полость, пп – проводящий пучок, пт – покровная ткань)
Все современные хвощи – многолетние корневищные травы с мутовками бурых редуцированных листьев, утративших хлорофилл (не следует путать с листьями мутовки зеленых веточек в узлах стеблей). Корневища у всех хвощей глубоко залегающие, сильно разветвленные, в их узлах часто закладываются почки. Почки на корневищах нередко образуют короткие утолщенные междоузлия (называемые клубнями), складывающиеся в короткие цепочки, богатые крахмалом. От узлов корневищ отходят многочисленные придаточные корни двух типов: растущие вниз, часто утолщенные, и направленные вверх или в стороны (рис. 3). Стебли прямостоячие или восходящие, простые или мутовчато-ветвистые, цилиндрические или с продольными ребрами, состоят из узлов и полых междоузлий. В узлах побегов находятся листовые влагалища, по верхнему краю которых расположены зубцевидные листовые пластины (рис. 4, В , Г ; 6). Листья хвощей срастаются основаниями в кольцо, сидящее на узле и прикрывающее растущую зону междоузлий, богатую сахаром. Верхушки листьев имеют форму узких зубцов. В почке эти зубцы сращены и образуют колпачок, который при росте стебля разрывается на верхушке. В дальнейшем при росте стебля в длину и толщину, листовые пластинки отходят друг от друга, но разрыв происходит иногда не по всем линиям срастания, поэтому число зубцов по краю влагалища может не соответствовать действительному числу листьев. Форма влагалища, цвет, форма и время жизни зубцов различны у разных видов, и эти признаки используются при определении хвощей.
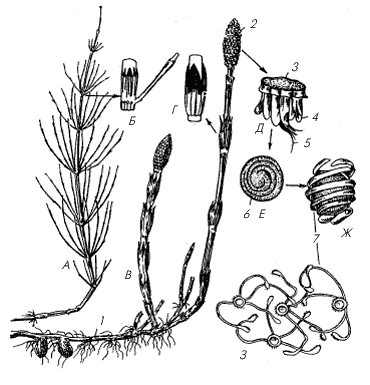
А – вегетативный побег; Б – спороносный побег; В, Г – листовые влагалища; Д – спорангиофор со спорангиями; Е–З – споры (1 – корневище с клубнями, 2 – стробил; 3 – щиток; 4 – спорангий; 5 – ножка; 6 – наружная оболочка споры, 7 – элатеры)
В основании листовых влагалищ хвощей закладываются почки, которые позже прорывают влагалища и образуют боковые побеги. Для возобновления растения особенно важны почки на тех узлах стеблей, которые находятся под землей, вблизи ее поверхности. Рядом с почкой закладывается корень.
По характеру надземных побегов хвощи можно разбить на две группы. У одних видов все надземные побеги однотипны. Они очень жесткие, обычно вечнозеленые (т.е. живут более одного сезона и в умеренной зоне перезимовывают) и развивают верхушечные стробилы.
У видов второй группы побеги бывают двух типов: одни – спороносные – буроватые или зеленые, а другие – вегетативные, зеленые. Побеги обоих типов появляются весной над поверхностью почвы, они нежнее по своей консистенции, чем побеги первой группы, и к зиме отмирают. Развитие вегетативных и спороносных побегов у видов второй группы идет по-разному. У хвоща полевого спороносные побеги появляются первыми ранней весной (рис. 4, В ). Они буровато-розовые, неветвистые, обычно толще быстро зеленеющих вегетативных побегов и после спороношения большей частью отмирают. У хвощей лесного и лугового спороносные побеги с хорошо развитыми стробилами появляются ранней весной одновременно с вегетативными. И те и другие вначале бледно-розовые, а затем зеленеют. После спороношения и отсыхания стробила спороносные побеги функционируют подобно вегетативным. Наконец, у таких видов, как хвощ болотный, хвощ приречный, хвощ боготский (E. bogotense – он обитает в тропиках Центральной и Южной Америки), спороносные побеги в момент их появления над поверхностью земли с трудом можно отличить от вегетативных – у них такая же зеленая окраска и сходные размеры.

Рис. 5. Формы листовых мутовок (а) и поперечные срезы побегов (б) хвощей:
1 – хвощ полевой; 2 – хвощ луговой; 3 – хвощ лесной; 4 – хвощ болотный; 5 – хвощ зимующий (а – листовое кольцо; б – поперечный разрез стебля)
Жесткость побегов хвощей объясняется тем, что в состав оболочки клеток эпидермы и, в меньшей степени, других тканей побега наряду с целлюлозой входит кремнезем. Частицы кремнезема располагаются в клеточной оболочке между тяжами целлюлозы, а на поверхности клеток эпидермы образуют сплошной и гомогенный, очень прочный слой с мелкими бугорками, приуроченными к выростам оболочки (рис. 5). Слой из кремнезема покрыт снаружи тонкой кутикулой с восковым налетом и играет не только механическую, но и защитную роль, так как благодаря ему хвощи практически не подвергаются нападению различных растительноядных моллюсков и насекомых.
Стробилы или спороносные колоски хвощей состоят из спорангиофоров, размещенных на общей оси. Они имеют вид шестиугольных щитков с ножками (рис. 4, Д ). На внутренней стороне такого щитка размещены 4–6 спорангиев. Когда находящиеся в них споры созревают, ось колоска немного удлиняется, и щитки спорангиофоров отходят друг от друга. Стенки созревших спорангиев лопаются, открывается продольная щель, и споры высыпаются. Лентовидные придатки (элатеры), образующиеся из наружной оболочки споры, способствуют сцеплению распространяющихся спор друг с другом (рис. 4, Ж , З ).
Споры могут прорастать только на затененной и влажной почве, где из них вырастает пластинчатый или ветвистый заросток. Заростки (гаметофиты) современных хвощей – очень маленькие, размером в несколько миллиметров, недолговечные зеленые растеньица.
Современные хвощи в отличие от вымерших каламитов – равноспоровые растения, на их заростках образуются органы (архегонии и антеридии), продуцирующие как женские (яйцеклетки), так и мужские (многожгутиковые сперматозоиды) половые клетки.
Сперматозоиды хвощей подвижны, поэтому оплодотворение происходит в воде – во время дождя или выпадения росы. Из зиготы развивается зародыш, а из него – без периода покоя – проросток, который через несколько месяцев превращается во взрослую особь нового бесполого поколения – спорофит (рис. 6).

Рис. 6. Чередование поколений в жизненном цикле хвощей (Equisetum arvense):
а – гаметофит (заросток); б – спорофит; в – спорангофор; г – спорангии; д – щиток спорангиофора; е – элатеры; ж – клубеньки
В природе хвощи обычно встречаются в виде клонов (групп растений, возникших путем вегетативного размножения), которые нередко занимают участки площадью в несколько десятков и даже сотен квадратных метров.
Хвощи отличаются крайним полиморфизмом. Например, у хвоща полевого, недавно поселившегося на хорошо освещенном участке с нарушенным растительным покровом, стебли обычно низкие, распростертые. Но позже, когда разовьется окружающая растительность, от более зрелых корневищ того же клона могут отходить мощные прямостоячие стебли с ветвистыми побегами. При приближении к северным границам ареала этого вида спороносные побеги, буроватые у растений умеренной зоны, сменяются зеленеющими, а потом – в тундре – и ветвистыми зелеными. Очень изменчивы бывают побеги и у других видов. Эти модификации, возникающие под влиянием многообразных факторов внешней среды (часто даже в пределах одного клона), наряду с многочисленными гибридами и наследуемыми уродствами, являются одной из причин разногласий между ботаниками в оценке объема рода.
Хвощи довольно широко распространены на Земле, особенно в Северном полушарии. Но в Австралии, Новой Зеландии и Тропической Африке аборигенных видов хвощей нет.
Встречаются хвощи в различных растительных сообществах, но обязательно с достаточным или избыточным увлажнением. По берегам водоемов, в сырых лесах, в некоторых типах низинных болот хвощи нередко доминируют в травянистом покрове.
Хвощи часто захватывают территории с нарушенным растительным покровом, образуя при этом чистые или почти чистые заросли в тех местах, где другие растения не могут жить, например, из-за обилия воды. Раз поселившись на какой-нибудь территории, хвощи, благодаря наличию глубоко залегающих корневищ, масса которых превышает массу надземных частей растения в несколько раз, успешно противостоят таким неблагоприятным воздействиям внешней среды, как засухи, лесные пожары и т.п., и успешно конкурируют с другими растениями, зачастую становясь широко распространенными и трудно искореняемыми сорняками пастбищ и полей.
Хвощи, растущие в условиях избыточного увлажнения, обладают признаками гигрофитов – у них слабо развита водопроводящая система, но хорошо представлена воздухоносная ткань. Одновременно эти растения могут нести и признаки ксерофитов – погруженные под поверхность эпидермы замыкающие клетки устьиц, закрытые устьица на старых участках стебля, сильно развитая механическая ткань, фотосинтезирующие стебли, редуцированные листья. Соотношение тех или иных признаков строения варьирует у разных видов.
В умеренной зоне России чаще всего можно встретить следующие виды хвощей.

Рис. 7. Хвощ луговой (Equisetum pratense Ehrh.)авт. рис. Шикина А.В.
Хвощ луговой (E.pratense, рис. 7) – многолетнее растение с удлиненным подземным черным корневищем без клубней. Спороносные побеги бледно-зеленые, простые, с воронковидными крупными влагалищами и свободными светло-коричневыми зубцами, с перепончатой каймой. Колосок длиной 0,8–2 см, цилиндрический. Спороношение в июне-июле. После спороношения, в начале лета, из спороносных побегов развиваются простые зеленые ветви, и они становятся похожими на вегетативные. Вегетативные побеги хвоща лугового имеют высоту 10–30 см и диаметр около 2 мм, они прямостоячие, также бледно-зеленые, внутри с большой центральной полостью и многими мелкими периферическими (рис. 5), с 8–14 шероховатыми ребрами, с простыми неветвящимися боковыми трехгранными веточками длиной 5–15 см, расположенными горизонтально или дугообразно книзу. Зубцы листовых мутовок широколанцетные, на ветвях яйцевидные, острые, в количестве 10–15 шт.
Продолжение следует
Хво-щи (Equisetum) - это многолетние травянистые растения, которые растут на влажных полях и лугах, на болотах и в сы-рых лесах. Хотя по внешнему виду они отличаются от папоротни-ков и плаунов , но по многим признакам схожи с ними. Хво-щи, как и папоротники , являются споровыми растениями . В настоящее время хвощи большой роли в образовании растительного покрова не играют. Хотя нередко хвощи обра-зуют заросли в тех местах, где другие растения существовать не могут.
Видовое разнообразие хвощей невелико — около 30 видов. В лесах на увлажненной поч-ве часто встречается хвощ лесной с сильно разветвленными поника-ющими боковыми веточками. На песчаных почвах и в оврагах рас-тет хвощ зимующий, на заболочен-ных местах, по берегам рек и озер произрастают хвощ болотный и хвощ приречный (рис. 88).
Хвощ полевой
Типичным представителем является хвощ полевой (рис. 87). Это многолетний сорняк, растущий на полях и пашнях. В почве находится ветвистое корневище с придаточными корнями и почками , из которых каждый год развивают-ся надземные побеги. При обработке почвы кусочки корне-вища хвоща не погибают, а из каждого вырастает самостоя-тельное растение. Поэтому с этим сорняком очень трудно бороться.
Строение
Хвощи имеют уникальные чле-нистые стебли. На стыках располо-жены листья. Стебель пропитан кремнезе-мом, который придает ему боль-шую прочность.
В благоприятных условиях споры хвоща, как и папоротника, про-растают в небольшие растеньица, непохожие на листостебельные. На них формируются органы полового размножения , в которых со-зревают половые клетки. При наличии капельной воды происходит оплодотворение. Из яйцеклетки образуется молодое растение хвоща с корневищем.
После образования спор весен-ние побеги отмирают, а от корне-вища отрастают зеленые летние побеги, похожие на маленькие со-сенки (см. рис. 87).
Стебли хвоща зимующего содержат значительное количе-ство кремнезема — твердого, хорошо полирующего вещества. Поэтому его стебли особенно жесткие, прочные. Они издав-на применялись для чистки металлической посуды и вместо наждачной бумаги.
Побеги некоторых хвощей (например, хвоща полевого) ис-пользуются в народной медицине как мочегонное и вяжущее средство.
Для хвощевидных характерно наличие побегов, состоящих из четко выраженных члеников (междоузлий) и узлов с мутовчато расположенными листьями. К хвощевидным принадлежат травянистые растения со стеблем длиной от нескольких сантиметров до нескольких метров. Древовидные формы, достигавшие высоты 15 м, а диаметра более 0,5 м, вымерли.
Высшие споровые растения - это первые наземные растения, обитающие во влажных, часто под пологом леса местах, или на болотах, или на полях с кислыми почвами. Господствовавшие в палеозое древовидные папоротники, хвощи и плауны в наше время представлены травами, за исключением тропических древовидных папоротников. Мхи же за этот период изменились мало, так как они занимают только свойственные им влажные места обитания. Этим растениям для размножения необходима вода, поскольку их гаметы - сперматозоиды - переносятся к яйцеклеткам только в капельно-жидкой воде, а заростки могут расти только на влажной почве. Жизнь в трудных условиях суши вела отбор на такие приспособительные признаки, как формирование вегетативных органов (корень, стебель, лист), органов размножения (архегонии, антеридии, спорангии), а также тканей. В цепях питания прошлых геологических эпох высшие споровые растения занимали ведущее место: они служили пищей для травоядных земноводных, пресмыкающихся. В настоящее время их роль как кормовых растений заметно уменьшилась, но значение в природе осталось: они удерживают воду в почве, создают условия для сохранения и прорастания семян голосеменных и покрытосеменных растений, являются средой обитания для животных. В хозяйстве человека велика роль древних древовидных форм, давших залежи каменного угля, который, как и торф, служит не только топливом, но и ценным химическим сырьем. Среди этой группы растений лишь хвощ полевой является трудноискоренимым сорняком полей с повышенной кислотностью почв. Высшие споровые - живые ископаемые, дошедшие до наших дней, поэтому их надо беречь и охранять. В Красную книгу СССР внесено 32 вида мхов, 6 видов папоротников; в Красной книге РСФСР содержатся 22 вида мхов, 10 - папоротников и 4 - плаунов. |

Характерной чертой хвощевидных является наличие у них своеобразных несущих спорангии структур - спорангиофоров. Подавляющее большинство хвощевидных - равноспоровые растения.
Размножение
Половое поколение - гаметофит (заросток). На гаметофитах образуются антеридии и архегонии. В антеридиях развиваются многожгутиковые сперматозоиды, а архегониях - яйцеклетки. Оплодотворение совершается при наличии капельно-жидкой воды, и из зиготы без периода покоя вырастает спорофит.
Классификация
Из хвощевидных ныне существует один класс хвощовые (или эквизетопсиды). Из семейства хвощовых наиболее распространен род хвощ. Хвощи часто составляют значительный процент травостоя лугов и заболоченных мест; распространены на кислой почве. Род объединяет около 30 видов; в нашей стране распространены 4, в Украине - 9. Наиболее часто, встречается у нас хвощ полевой, хвощ луговой, хвощ болотный, хвощ топяной и хвощ лесной.
Хвощ
Травянистое растение, имеющее однолетние надземные побеги. Небольшое количество видов вечнозеленые. Размеры стеблей хвощей сильно варьируют: встречается карликовые растения со стеблем высотой 5-15 см и диаметром 0,5-1 мм и растения со стеблем длиной в несколько метров (у хвоща многощетинкового стебель достигает длины 9 м). Хвощи тропических лесов достигают высоты 12 м. Подземная часть - корневище, ползущее, ветвистое, в котором могут откладываться питательные вещества (образуются клубни) и которое служит органом вегетативного размножения. Надземные побеги нарастают верхушкой. Летние побеги - вегетативные, ветвистые, ассимилирующие, состоят из члеников, с хорошо развитыми междоузлиями. От узлов ответвляются мутовчато расположенные и также расчлененные ветви. Листья малозаметные, срастаются в зубчатые влагалища, которые одевают нижнюю часть междоузлия. В клетках эпидермы стебля часто откладывается кремнезем, поэтому хвощи - плохой корм.
Весенние побеги - спороносные, неассимилирующие, неветвистые, на их верхушке образуются спороносные колоски. После созревания спор побеги отмирают. Споры шаровидные, с четырьмя пружинистыми лентами, зеленоватые, прорастают в заростки, однополые - мужские или женские. Бывают случаи, когда антеридии и архегонии зозникают на одном заростке. Из оплодотворенной яйцеклетки вырастает предросток, а затем - взрослый хвощ.
Хвощевидные Equisetophyta(sphenophyta).
Эквисетофита - от латинского конская щетина. Сфенофита - название членистые отражает особенности строения побегов. В узлах находится интеркалярная меристема.
Хвощевидные как и плауновые появились в верхнем девоне. Палеозойская эра, наибольшее распространение имели в позднем палеозое(каменноугольный период) входили в состав обширных лесов, были крупными,древовидными, высокими. На сегодняшний день наблюдается последние немногие остатки некогда обширной группы вымершей в начале мезозоя.
Современные представители, это многолетние растения, корневищные, травянистые,все относятся к одному роду хвощ, преобладающее поколение в цикле – это спорофит, тело спорофита разделено на хорошо развитый надземный побег. Высота его от нескольких сантиметров до нескольких метров. Так же имеется хорошо развитое корневище, от него отходят придаточные корни.
Все хвощевидные характеризуются четким разделением стебля, боковых ветвей и корневища, на узлы и полые междоузлия. Ветвление мутовчатое.
У хвоща полевого формируются стебли двух типов
1.Безхлорофильные, несущие споры, быстро отмирающие.
2.Зеленые, вегетативные.
(Те и другие продольно бороздчатые.)
У других хвощей например Хвоща лесного, стробил развивается на вегетативном побеге.
Листья современных хвощовых очень мелкие, имеют вид чешуек или зубцов, но происхождение у них отличное от листьев у плаунов. Лист макрофильной линии эволюции.
У вымерших хвощевидных из класса клинолистные, листья были относительно крупные. Имели дихотомическое жилкование, их эволюционное развитие заключалось в утолщении, сближении, срастании, редукции.
В связи с тем что листья у современных хвощей маленькие функцию листьев взял на себя стебель.
Корни только придаточные, отходят от корневища.
Спороносный колосок(стробил)
У вымерших хвоще стробилы имели довольно сложное строение, у современных более простое: на оси колоска мутовчато располагаются спорангиефоры(спорофиллы,листья несущие спорангии) они резко отличаются от листьев, лишены хлорофилла и несут спорангии. Хвощи равноспоровые.
Среди ныне живущих растений нет разноспоровых, у вымерших хвощей разноспоровость была. У клинолистов, у каломитов. Споры у хвощей отличаются от иных спор у высших растений: у низ 3 оболочки, 2 из них экзина и интина. Имеется 3я перина(наружная). Которая разделяется на 2 лентовидные пружинки элотеры (обладающие способностью к движению). В зависимости от влажности окружающей среды они сворачиваются вокруг тела споры или во влажую погоду наоборот раскручиваются разрыхляются и образуется рыхлая споровая масса, которая может вылетать из спорангия. Эти споры выпадают из спорангия и прорастают в непосредственной близости от спорангия. Гаметофиты современных хвощовых имеют вид лопастной или изрезанной верхней части зеленой пластины в несколько миллиметров толщиной. Многослойные, крепятся к почве с помощью ризоидов. Долгое время гаметофиты считали раздельнополыми, хотя все современные хвощи равноспоровые. В результате искусственного выращивания и наблюдения определили, что гаметофиты потенциально обоеполые, но развитие начинают как мужские и женские. На женских с возрастом могут образоваться антеридии, на мужских архегонии не образуются никогда, таким образом у хвощей наблюдается морфологическая равноспоровать и физическая разноспоровость.
Классификация
Отдел хвощевидные
1.Класс клинолистные(обладали вторичным ростом, образовывали заросли, длинна метр, поверхность ребристая, причем ребра переходили из одного междоузлия в другое без чередования, на стеблях мутовки листьев.У наиболее древних рассченные листья, у поздних клиновидная пластина, стебли сложного анатомического строения, в центре первичная ксилема(прото и мето ксилема) между ее лучами флоэма, между ними камбий, в наружных слоях феллоген образовывающий пробку.10 см в длину. Большинство клинолистов равноспоровые, некоторые разноспоровые)
2.Класс Хвощевые
А)Порядок Каламитовые(вымерли, появились в девоне, наибольшее развитие в карбоне и исчезают в юрском периоде.Эти деревья хвощи. Обитали в болотной почве из которой вызодили побеги высотой до 30 метров, стебли имели ущлы и междоузлия, ребра соседних узлов чередовались. От корневищ многочисленные придаточные корни. Листья линейные с одной жилкой. Среди них были как равноспоровые так и разноспоровые)
Б)Порядок Хвощевые(представлен одним семейством, одним родом, в Африке как заносные растения, несмотря на примитивное строение и древнее происхождение успешно конкурирует с наземными растениями. Циклы развития
Ископаемые подобного рода встречаются чрезвычайно редко, это единичные экземпляр. Здесь изображен паук, который атакует свою жертву. Потом и одного и второго накрыла смола хвойного дерева, в результате мы получили кусок янтаря с яркой картиной жизни членистоногих далекого, очень далекого, прошлого.
Если подумать, то вероятность получения подобной сценки в куске янтаря исчезающе мала. Тем не менее, счастливая случайность предоставила ученым, да и обывателям, возможность оценить разнообразие жизни 100-миллионолетней давности.
Ископаемый носорог (10 миллионов лет назад)
Череп большого носоорога был найден в Турции. Это животное погибло случайно, а причина - извержение вулкана, случившееся 10 миллионов лет назад. Палеонтологи считают, что вулканические бомбы - главная причина смерти животного, которое было просто забито до смерти падающими с неба раскаленными кусками породы. Череп буквально «запекся», и это не удивительно, поскольку температура пород достигала 450 градусов Цельсия.
По словам специалистов, останки млекопитающих очень редко встречаются в вулканических породах, по большей части потому, что высокая температура, актуальная при извержении вулкана, уничтожает зачастую даже кости. Но в данном случае ученым повезло, и череп носорога был найден практически неповрежденным.
Мегалодон (ископаемая гигантская акула)

Акула, которой принадлежала гигантская челюсть, изображенная на фото, погибла около полутора миллионов лет назад. Размер этой акулы составлял 16 метров, и, как видим, человек был бы вовсе неинтересен такой громадине в качестве добычи. Вероятно, мегалодоны охотились на более крупных существ, которых в то время было в море очень много. И да, на этой челюсти насчитали 182 зуба, можете себе представить? В наше время практически нет морских животных, которые могли бы сопротивляться мегалодону.
Гигантская пчела (50 миллионов лет назад)

Ископаемые насекомые, которых порой находят палеонтологи, имели весьма примечательные размеры. В качестве примера можно привести громадную пчелу, найденную в осадочных породах (вернее, найден ее отпечаток). Ее тело имело длину свыше пяти сантиметров. Представляете, как выглядел рой таких пчел? Наверное, их боялись даже динозавры, причем не только мелкие или средние.
Для сравнения показано тело колибри, которое примерно равно длине тела пчелы (Если не учитывать перья хвоста). Сейчас такого размера достигают только матки пчел некоторых видов, которые водятся в тропической Африке.
Гигантский пингвин (35 миллионов лет назад)

Останки (окаменевшие, само собо) гигантского пингвинча были найдены в Перу, о чем была написана крупная научная статья. И даже, если не вдаваться в науку, то птичка была действительно примечательной. Масса ее составляла 54-59 килограмма, а размеры в два раза превышали размер императорского пингвинча, который считается крупнейшим видом пингвинов на данный момент.
У пингвина был удлиненный клюв, которым, как полагают ученые, было удобно ловить рыбу в экваториальном море того времени.
Саблезубая белка (100 миллионов лет)

Да, представляете, не только тигры были саблезубыми много лет назад. Были и белки, которые тоже имели зубы в виде небольших забель. И размер этих зубов составлял около пятой части размера тела белки. Жили эти животные примерно 100 миллионов лет назад, намного раньше саблезубых тигров.
По словам палеонтологов, найти такой редкий вид млекопитающего, да еще такого древнего - нечто вроде выигрыша в лотерею. По мнению ученых, внешний вид этой «белочки» вполне соответствовал внешнему виду мультяшного героя в «Ледниковом периоде».
Ископаемый кит

Окаменевший скелет кита, выступающий из осадочных пород дна океана, был обнаружен не учеными, а серферами, около калифорнийского побережья. В момент обнаружения случился сильный отлив, и обнажившееся дно показало изумленным спортсменам-любителям большой скелет какого-то животного.
После того, как фотографии скелета попали в Сеть, нашлись ученые, которые определили окаменелость, как останки ископаемого кита времен Плиоцена.
Гнездо динозавров (70 миллионов лет)

В Монголии один из палеонтологов случайно открыл целое гнездо динозавров, с останками 15 динозавров-малышей. Сохранились они очень хорошо, и ученый смог детально проанализировать находку. В общем-то, одним из результатов исследования как раз и явилось доказательство «гнезования» динозавров, которые, как оказалось, первые недели после вылупления из яйца проводили в гнезде. Так что у динозавров был вполне себе приличный материнский инстинкт, благодаря которому и выкармливались малыши.
Может быть, за ними ухаживал и «отец», но это, конечно же, доказать сейчас практически невозможно. Что явилось причиной гибели динозавриков, пока неясно.
Хвощ (300 миллионов лет)

Это не очень редкая находка, но настолько хорошо сохранившийся хвощ, что отпечатались даже неровности «листьев» (у хвоща таковых нет), в общем-то, является событием примечательным.
Странный червяк (542 миллиона лет назад)

Окаменелые останки червя, который жил пол-миллиарда лет назад, были найден учеными в канадском Национальном Парке Yoho. У червей, как известно, вовсе нет скелета, и поэтому находки, подобные этой, встречаются крайне редко.
Палеонтологи считают, что этот червь является промежуточнм звеном развития между, собственно, червями, и остальными обитателями морей и океанов. Т.е., это одно из ранее ненайденных ключевых «звеньев» эволюции.
Имя: Антон Силуанов (Anton Siluanov) День рождения: 12 апреля 1963 (54 года) Место рождения: г. Москва Вес: 85 кг Рост: 178 см Знак Зодиака: Овен Восточный гороскоп: Кролик Деятельность: политик, министр финансов РФ Биография Антона СилуановаАнтон Герман
В закрепите одинаковый порядок учета транспортно-заготовительных расходов (ТЗР) как по товарам, так и по материалам. Это позволит вам избежать пересчетов, если придется использовать в качестве , ранее учтенные как товары. А если вы выберете вариант учета,
Бюджетная классификация Российской Федерации является группировкой доходов, расходов, источников финансирования дефицитов бюджетов и операций публично-правовых образований в бюджетной системе. Она применяется для составления и исполнения бюджетов, формиро






